The reference is: Benis AM and Hobgood DK. Dopamine receptor DRD3 codes for trait aggression as Mendelian recessive. Medical Hypotheses 2011:77(5) (In press).
A preprint of the paper may be accessed here.
Haplotype associated with protection from schizophrenia leads to candidate gene associated with trait aggression in NPA theory
Just as consideration of the association of autism and OCD (obsessive-compulsive disorder) was helpful in positing a candidate gene for perfectionism (trait P), we believe that schizophrenia (SZ) can be helpful in deducing which genes are associated with traits A and N.
To summarize, NPA theory posits three genetic traits that determine genetic character type:
sanguinity, or narcissism, denoted by N
perfectionism, denoted by P
aggression, denoted by A
The genes for trait A and for trait N are considered to be complementary, as either trait A or trait N (or both) must be present in any individual. Thus, traits A and N may be present separately or together, but the absence of both would be lethal to a fetus and result in miscarriage or stillbirth.
The presence of both traits A and N, fully expressed, appears only in the character types dominant NA and dominant NPA.
According to NPA theory, a basic predisposition to SZ is dependent on the presence, or absence, of the traits N and A (Benis, 1990). The character types most vulnerable are those having either trait A or trait N, but not both. Thus, having both traits A and N, fully expressed as dominant NA or NPA character type, is a phenotypic marker for low vulnerability to SZ. This has ramifications at the level of 1) an individual, 2) a family pedigree, and 3) a population.
At the level, of an individual this means that an NA or NPA dominant character type would have low vulnerability to SZ. Lack of having both traits would indicate that the individual is not in the group of low vulnerability, but not necessarily in a subgroup of high vulnerability.
At the level of a family, the presence of both traits A and N throughout the family means that the family has low vulnerability to SZ. Again, lack of having both traits would mean that the family is not in the group of low vulnerability, but not necessarily in a subgroup of high vulnerability.
At the level of a population, the presence of large numbers of individuals with both traits A and N would indicate a large number of individuals having protection from SZ. In a population having a high frequency of N trait (most areas of the world), the presence of the A trait would appear as marker associated with protection from schizophrenia.
We refer now to the landmark paper of Costas and colleagues (2009) A common haplotype of DRD3 affected by recent positive selection is associated with protection from schizophrenia.
Costas et al. found evidence for a haplotype of gene DRD3 (dopamine receptor) on chromosome 3 being associated with protection against SZ. The haplotype was found to have altered in frequency due to natural selection, mostly after the Out of Africa recent migrations of modern man. The haplotype was found to be of low frequency in indigenous sub-Saharan African groups, but also in Polynesia.
Below is the relevant figure from the paper by Costas et al., where the numbers refer to sample size and the dark areas of the circles denote frequency of the DRD3 haplotype.
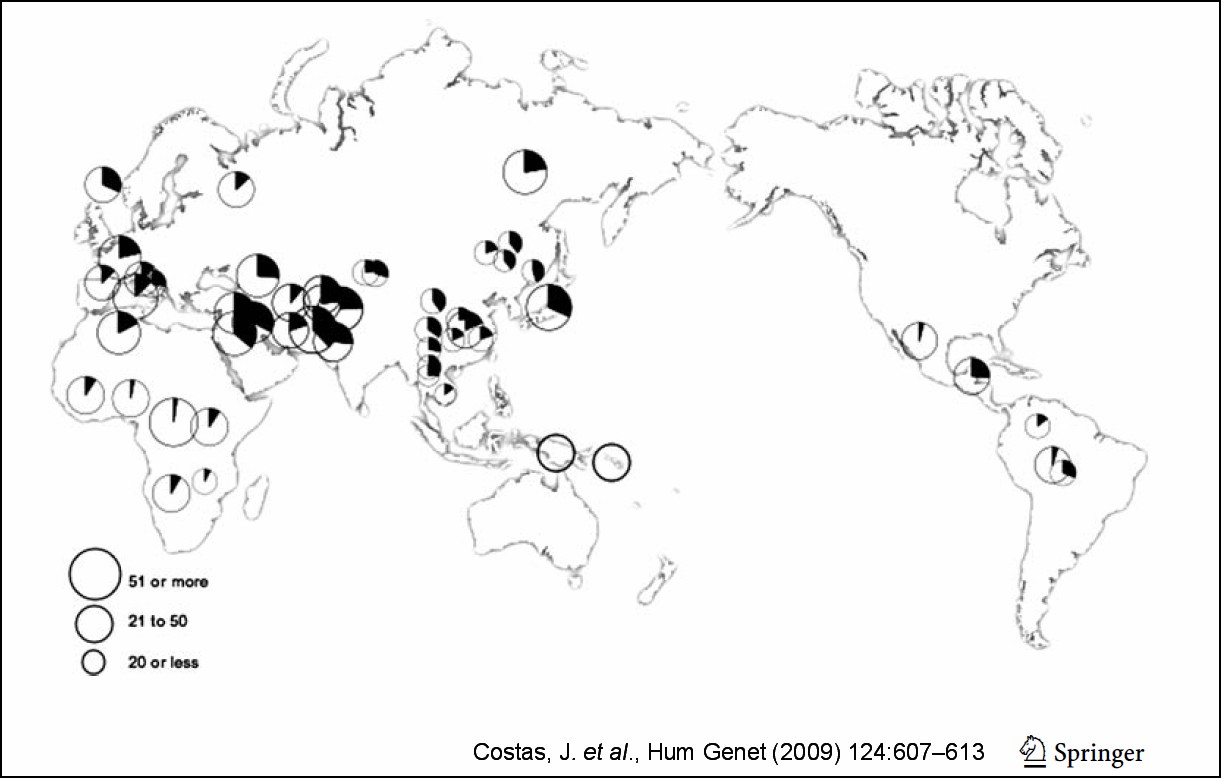
What is interesting is that in NPA theory, we had deduced that the trait A (trait aggression) was of low frequency in Sub-Saharan Africa and Polynesia, with the highest frequency in the Mideast and intermediate frequency in central Europe.
The map below is our Habitancy map of NPA character types. It is based on a variety of sources from the anthropological literature, personal accounts, popular press, as well as current images and videos. It is admittedly approximate and incomplete.
Nevertheless, the main findings shown on the Habitancy Map are:
1) the sanguine N trait is almost omnipresent in the world. The only major regions where the sanguine N trait was found to be of low frequency was Eastern Europe (e.g., Poland and extending into Russia); the Balkans (e.g., Serbia) and Aboriginal Australia.
2) the A trait is of low frequency in most of Sub-Saharan Africa and is highest in the Mideast. It is also of very low frequency in Polynesia.
We had also concluded (Benis, 2008) that the A trait was highly affected by natural selection after the Out of Africa migrations, resulting in dramatically different frequency of the A trait according to geographical location.
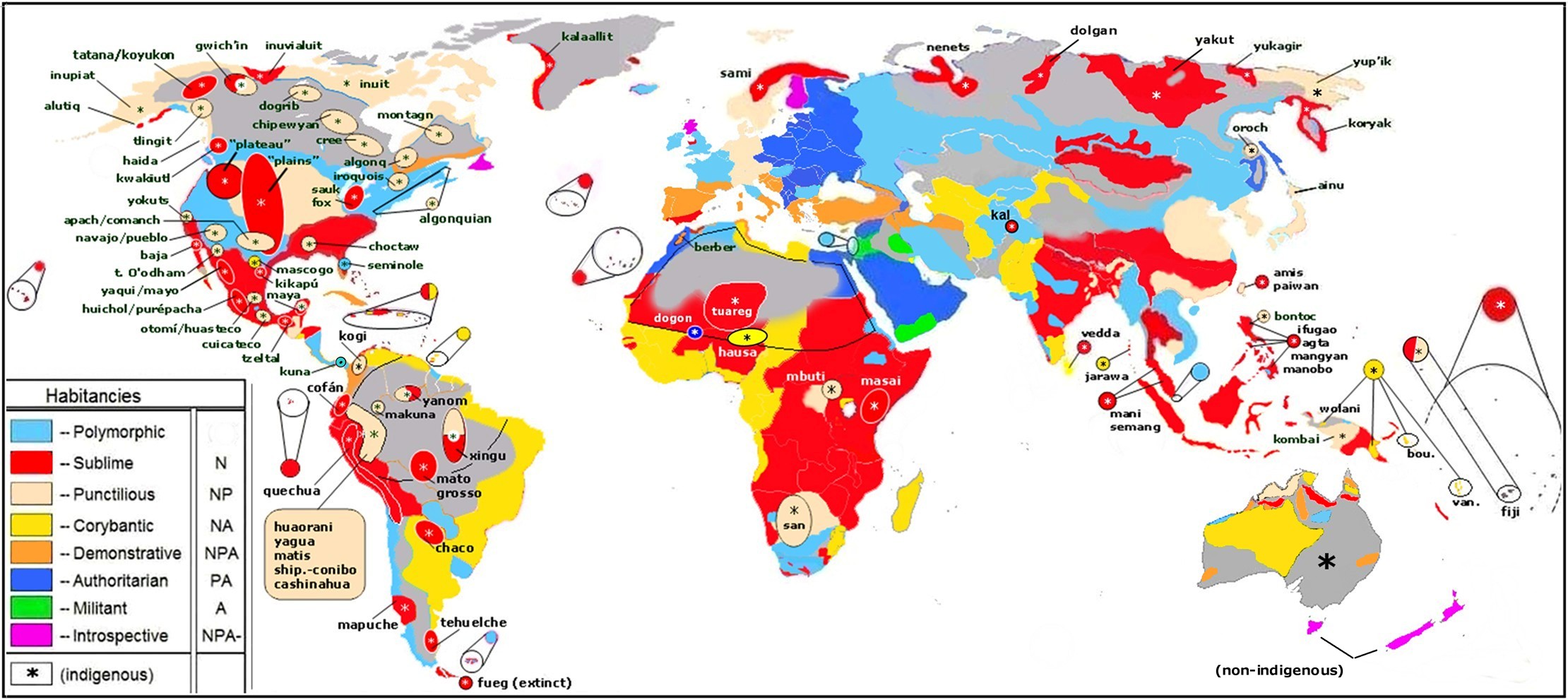
N.B. The color RED denotes areas where the sanguine trait N is present with little trait A (trait aggression).
In NPA theory, given the widespread prevalence of the N trait, the increase of the A trait in a local population would lead to increased frequency of individuals (NA and NPA dominant types) having protection from SZ. The A trait would appear as phenotypic marker for protection from SZ.
From the above, we posit that that the DRD3 haplotype found by Costas et al. is associated with trait aggression, or trait A in the NPA model.
In the paper of Costas et al. it appears that the protection from schizophrenia is relevant in the statistical sense of individuals in a population having lower vulnerability. It is not clear whether an individual homozygous for the protective haplotype would have lower vulnerability, or whether the vulnerability is different in heterozygous and homozygous individuals. Clearly, family studies are necessary.
In NPA theory, the mode of inheritance of trait A is not yet clear and is not likely to be simple Mendelian involving only one gene. In some family pedigrees, inheritance of trait A was most consistent with an autosomal recessive mechanism. However, it is possible that several genes (perhaps the X-linked MAO-A) are involved. It would be premature, then, to hypothesize how the DRD3 haplotype is associated with trait A. It could be that DRD3 is involved directly in neurotransmitter function or that the haplotype found by Costas et al. is a marker by genetic linkage or otherwise. It could be that the haplotype would have to be present in the homozygous state to offer protection from SZ.
A reasonable first study would be to test if healthy individuals homozygous for the DRD3 haplotype score higher in trait aggression than control subjects in a personality inventory.
Finally, our hypothesis above includes the supposition that the A trait is a protection against SZ only in a population where the N trait is of high prevalence. Therefore, having the DRD3 haplotype would not be protective in a (non-sanguine) area where the N trait has low prevalence and the A trait is of high prevalence. Thus, we predict that in relatively non-sanguine areas like Serbia, Poland, Russia or the Australian Aboriginal population (where SZ is likely to be found in the A and PA character types) the frequency of the DRD3 haplotype will be found to be relatively high in those groups of patients with SZ.
Acknowledgement: Donna K. Hobgood, M.D. began the inquiry of the relevance of neurotransmitter function in NPA theory and also contributed to the Habitancy Map shown above.
____________________
REFERENCES:
Costas, Javier Noa Carrera Eduardo Domínguez Elisabet Vilella Lourdes Martorell Joaquín Valero Alfonso Gutiérrez-Zotes Antonio Labad Ángel Carracedo, Human Genetics (2009) 124:607613: A common haplotype of DRD3 affected by recent positive selection is associated with protection from schizophrenia.
Benis A.M. (1990), Speculations in Science & Technology (1990) 13:167-175. A theory of personality traits leads to a genetic model for borderline types and schizophrenia.
Benis A.M. (2008), NPA Theory of Personality, Internet Edition, originally published by Psychological Dimensions, Inc., New York, 1985.
__________________
To comment on this posting, please refer to the thread below, or start a new thread.